 Kolmann Lab
Form.Function.Evolution
Kolmann Lab
Form.Function.Evolution
Biomechanics & Scaling
Think about it: somehow your genetic code allows growth to proceed at an astonishing rate in many animals, with all the individual structures, organs, and physiological mechanisms proceeding apace. The ability of the body to function within a changing paradigm (nature) AS a changing paradigm is both a wonder in itself and probably the reason why life is so hardy in the first place. The intertwining of physical laws and natural selection, how the two interact and become at odds, is the primary interest of biomechanics. The study of how physical laws effect the growth and development of organisms, particularly in different environments, deals with scaling: differential growth and development of structures and modules within an organism throughout its life.
Feeding kinematics & insectivory in freshwater stingrays
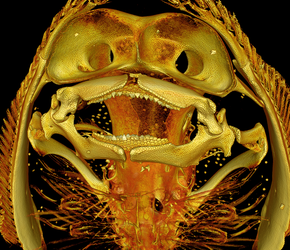
Despite considerable research into the evolution of feeding in fishes, little attention has focused on aquatic insectivory, especially in elasmobranch fishes. Many freshwater stingrays prey on aquatic insects, primarily chironomid larvae, but also orthopteran, dipteran, and odonate larvae. Stingrays are known for lengthy prey-processing behaviors, but the manner in which they can successfully disarticulate and comminute insect prey is poorly understood.
The Potamotrygonidae are an obligatory, marine-derived family of stingrays found in the Amazon, Orinoco, and Guiana Shield drainages of South America. Within this family, at least five species feed on insect or benthic invertebrates, including the widespread species Potamotrygon motoro. The flexible, unfused mandibular symphysis in potamotrygonids may allow for tooth reorientation (as documented in electric rays by Mason Dean). In addition, the highly decoupled cranial skeleton of these rays, in which the jaws suspended from the cranium via two intermediate skeletal elements, the paired, bilateral hyomandibular and angular cartilages, permits a jaw protrusion mechanism which is highly kinetic. These morphological traits, along with the behavioral ability of potamotrygonids to repeatedly move prey in and out of the oral cavity presumably contribute to the ability of stingrays to effectively macerate insect prey.
Kolmann, M.A., Welch, K.C., Summers, A.P., Lovejoy, N.R. (2016). Always chew your food: freshwater
stingrays use mastication to process tough insect prey. Proceedings of the Royal Society: Part B.
The fitness advantage of higher feeding performance: a case study using two populations of long-lived, durophagous stingrays.

Feeding performance scales in most vertebrates in relation to size: as animals get bigger, so do many of their performance capabilities. Higher performance translates into greater foraging success and therefore, greater fitness in larger animals. But what about in animals which show differential growth between skeletal structures and muscle performance? How might sympatric species distance themselves with regards to resource partitioning? By changing size, physiology, or both? Cownose rays have been singled out leading to declines in harvestable shellfish stocks - but are all these Rhinoptera performing in a similar manner?
Objectives: 1) How does the feeding performance of Virginia cownose rays compare to their Florida relatives? 2) Does size versus physiology mediate these differences in performance? 3) What can these experimental scenarios tell us about the effects these rays may have on harvestable shellfish populations?
Kolmann, M.A., Huber, D.R., Fisher, R., Grubbs, R.D. (2018). Alternative muscle scaling patterns explain increased foraging performance between two species of durophagous stingrays.
Ontogeny of feeding performance in cownose rays
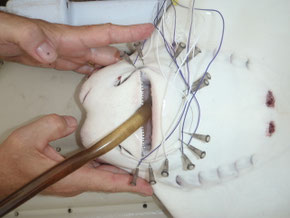
The cownose ray is a source of controversy and increasing media attention due to interactions with commercial fisheries and on-bottom aquaculture operations for bivalves such as oysters, clams, and bay scallops. Claims of increases in the population of cownose rays coupled with increases in damage to shellfish culture, grow-out, and restoration operations have led to the development of commercial harvesting for cownose rays in regions such as Chesapeake Bay, though no stock assessments have been completed and no management plans exist. We seek to empirically evaluate the jaw biomechanics of the cownose ray relative to the mechanical properties of commercial bivalves in order to inform management and stakeholders and perhaps suggest alternative means of mitigating interactions.
Objectives: 1) construct a model which generates maximal bite forces from cranial anatomy arrangement in Rhinoptera bonasus, 2) validate biomechanical model using in-vivo measurements of stimulated muscle contraction, 3) determine if scaling patterns of feeding performance over ontogeny are consistent with other durophagous fishes, 4) at what sizes are commercial and non-commercial shellfish at risk from predation across the ontogenetic trajectory of Rhinoptera?
Kolmann, M.A., Huber, D.R., Motta, P.J. and Grubbs, D.R. (2015). Ontogeny of bite force generation in a durophagous stingray, Rhinoptera bonasus. Journal of Anatomy.
Decoupling jaw morphology from performance: prey-crushing in durophagous stingrays.

Durophagy has evolved several times within the elasmobranch fishes. The stingray family Myliobatidae is composed almost entirely of durophagous taxa, except for the planktivorous manta and devil rays. These durophagous rays use rigid jaws, pavement-like dentition, and hypertrophied jaw muscles to crush and consume mollusks and crustaceans. The curvature of the upper and lower tooth plates vary considerably from one another, as well as between related taxa. We hypothesize that this difference in curvature will affects feeding performance and, therefore, patterns of ecological niche partitioning. Using aluminum stingray jaw replicas fixed to a mechanical loading frame, we measured the forces necessary to fracture uniformly manufactured ceramic tubes, live gastropods (Nucella sp.), live bivalves (Nuttalia, Mytilus), and 3D-printed shell replicas.
Kolmann, M.A., Crofts, S.A., Summers, A.P., & Lovejoy, N.R. (2015). Decoupling jaw morphology from performance. Journal of Experimental Biology.
Constraints on the feeding ecology of juvenile horn sharks
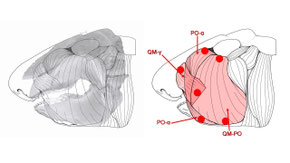
Organismal performance changes over ontogeny as the musculoskeletal systems underlying animal behavior grow in relative size and shape. As performance is a determinant of feeding ecology, ontogenetic changes in the former can influence the latter. The horn shark, Heterodontus francisci, consumes hard-shelled benthic invertebrates, which may be problematic for younger animals with lower performance capacities. Scaling of feeding biomechanics was investigated in H. francisci (n = 16, 19–59 cm standard length (SL)) to determine the biomechanical basis of allometric changes in feeding performance and whether this performance capacity constrains hard-prey consumption over ontogeny. Positive allometry of anterior (8–163 N) and posterior (15–382 N) theoretical bite force was attributed to positive allometry of cross-sectional area in two jaw adducting muscles and mechanical advantage at the posterior bite point (0.79-1.26). Mechanical advantage for anterior biting scaled isometrically (0.52). Fracture forces for purple sea urchins Strongylocentrotus purpuratus consumed by H. francisci ranged from 24 to 430 N. Comparison of these fracture forces to the bite force of H. francisci suggests that H. francisci is unable to consume hard prey early in its life history, but can consume the majority of S. purpuratus by the time it reaches maximum size. Despite this constraint, positive allometry of biting performance appears to facilitate an earlier entry into the durophagous niche than would an isometric ontogenetic trajectory. The posterior gape of H. francisci is significantly smaller than the urchins capable of being crushed by its posterior bite force. Thus, the high posterior bite forces of H. francisci cannot be fully utilized while consuming prey of similar toughness and size to S. purpuratus, and its potential trophic niche is primarily determined by anterior biting capacity.
Kolmann, MA, Huber, DR. 2009. Scaling of feeding biomechanics in the horn shark Heterodontus francisci: Ontogenetic constraints on durophagy. Zoology. 112:351–361.